Qual a Importância da Anidrase Carbonica Para o Sistema Tampão Sanguíneo?
Exclusivo
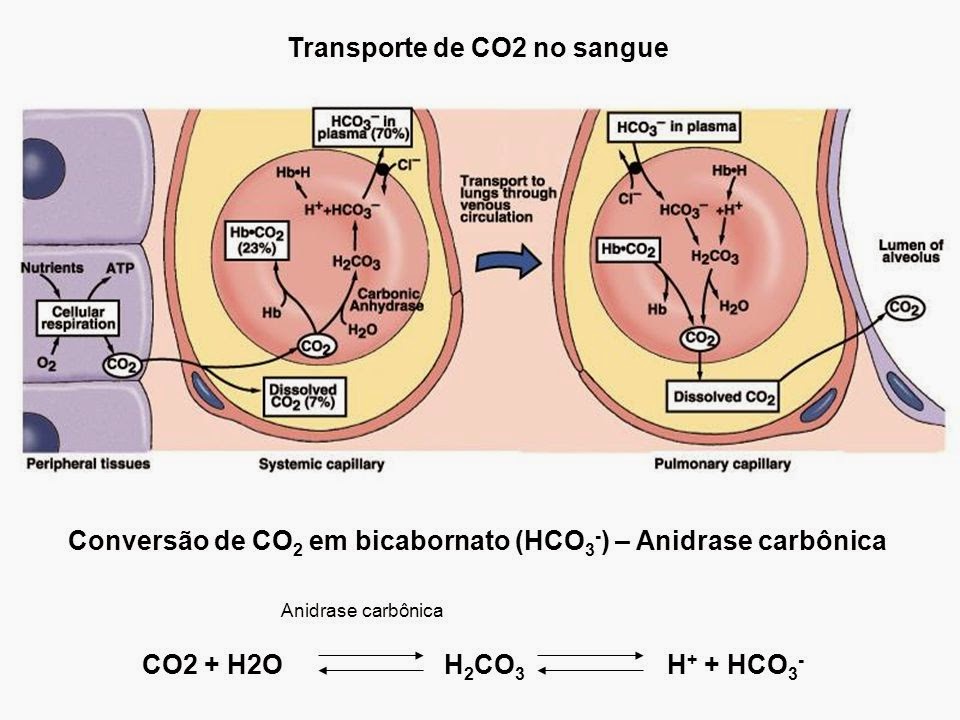
O CO2 produzido no interior do músculo esquelético precisa sair finalmente do corpo por meio de ventilação pelo pulmão. Para chegar, o CO2 se difunde do espaço intracelular para o sangue médio do veículo convectivo com os dois compartimentos, plasma e eritrócitos.
Dentro do corpo, o CO2 é enviado em três estruturas distintas: genuinamente desintegrado, como HCO3-, ou como carbamato. O compromisso geral destas três estruturas com o veículo em geral está mudando ao longo deste caminho final. Por conseguinte, a energia da troca deve ser considerada.

Ilustração do CO2
A anidrase carbónica acelera a resposta de hidratação/parqueação entre CO2, HCO3-, e H+. No músculo esquelético, diferentes isozimas de anidrase carbónica são restringidas no interior dos eritócitos, mas, ao mesmo tempo, estão ligadas ao divisor estreito, desta forma aberto ao plasma; ligadas ao sarcolema, proporcionando assim ação sinérgica dentro do espaço intersticial; e ligadas ao retículo sarcoplásmico.
Em alguns tipos de fibras, a anidrase carbónica está adicionalmente presente no sarcoplasma. Na prática do músculo esquelético, o corrosivo láctico contribui com medidas colossais de H+ e por estas influencia o compromisso geral dos três tipos de CO2.
Com um modelo hipotético, a associação espantosa de respostas e formas de transporte envolvidas com o comércio de CO2 foi quebrada.
Apresentação
Uma das necessidades significativas do corpo é matar o CO2. A enorme, embora excepcionalmente importante, medida de CO2 que é entregue dentro das células musculares precisa finalmente deixar o corpo por meio da ventilação do espaço alveolar.
Para chegar, a disseminação do CO2 precisa acontecer do espaço intracelular dos músculos para o sangue médio do veículo convectivo, e a dispersão para fora do sangue precisa ocorrer no espaço gasoso pulmonar sobre a obstrução alveolocapilar.
O dióxido de carbono no corpo está disponível em três estruturas únicas: quebrado, ligado como bicarbonato, ou ligado como carbamato.
O compromisso geral destas várias estruturas com o transporte de CO2 e de grandes dimensões muda exclusivamente ao longo deste percurso final, à luz do facto de que, para a disseminação através de obstáculos de camadas, outra estrutura é mais adequada do que para o transporte dentro de compartimentos intra ou extracelulares.
Desta forma, a energia do comércio entre as estruturas torna-se basicamente significativa. Além disso, os resultados de uma dessas trocas, a resposta de hidratação de CO2, HCO3-, e H+, são necessários para um sortimento extraordinário de outras capacidades celulares, por exemplo, descarga de corrosivos ou base e algumas respostas de digestão mediadora. Na prática do músculo esquelético, o outro “resultado final” da digestão, o corrosivo láctico, contribui imensas medidas de H+ e por estas influencia a transcendência dos três tipos de CO2, com base no facto de que o HCO3- tal como o carbamato, dependem basicamente da centralização do H+.
A conversa sobre o veículo geral do CO2 no músculo esquelético precisa considerar este compromisso de corrosivo láctico e sua inclusão na energia e equilíbrio das reações de CO2. Esta dependência do CO2 e da extremidade corrosiva láctica é uma parte significativa desta auditoria, que na medida em que sabemos não foi inspecionada antes em detalhe.
Transporte De Dióxido De Carbono No Sangue
Formas de Transporte de CO2 no Sangue
As formas de transporte de dióxido de carbono no sangue foram totalmente examinadas por Klocke. Nós apenas esboçamos rapidamente seu compromisso individual com o comércio de CO2 em geral.
CO2 no Sangue
CO2 Dissolvido
Apenas uma pequena parte, ∼5% de todo o conteúdo dos vasos sanguíneos, está disponível como CO2 decomposto. Utilizando um coeficiente de dissolvabilidade Sco2 de 3,21 × 10-5M/Torr (35) para plasma a 37°C, isso dá 1,28 mM de CO2 decomposto, ou, utilizando Sco2 de 3,08 × 10-5 M/Torr (7), 1,23 mM de CO2 desintegrado a um Pco2 de 5,32 kPa (40 Torr).
Muito quieto, o compromisso de dividir o CO2 com a distinção do foco arteriovenoso absoluto do CO2 é de apenas 8% em ∼10. Em qualquer caso, durante um exercício substancial, o compromisso de decompor o CO2 pode se expandir sete vezes e, em seguida, perfazer cerca de 33% do comércio total de CO2.
Co2 Ligado Como Hco3-
A maior parte do CO2 em todos os compartimentos está ligada como HCO3-. A proporção de HCO3- sobre o CO2 decomposto é dada pela condição Henderson-Hasselbalch:
https://www.youtube.com/watch?v=0j_ais_ogw8
pH=pKa′+log[HCO3-]SCO2×PCO2
O pKa′ tem uma estimativa típica de 6,10 no plasma humano a 37°C e flutua com a temperatura e qualidade iônica. Tem todas as marcas de ser marginalmente extraordinário em soro e plaquetas vermelhas: soro, 6,11; eritrócitos oxigenados, 6,10; eritrócitos desoxigenados, 6,12.
Durante uma carga substancial e excepcional do músculo, níveis elevados de corrosivos lácticos estão disponíveis, apesar do CO2, irritando a queda do pH. Com este pH baixo, a porção de HCO3- em CO2 absoluto é diminuída.
Apesar do fato de que no pH 7,4 HCO3- é 20-sobreposição contrastada e quebrada de CO2, é apenas 13-sobreposição com o típico pH de 7,2, e a proporção pode cair para um monte de estima mais baixa em estimativas de pH plasmático de significativamente abaixo de 7 durante o exercício máximo.
Desta forma, apesar de inquestionavelmente a distinção arteriovenosa ser maior durante o exercício do que durante o descanso, o compromisso geral do HCO3- com o comércio em geral é menor. Para o caso da atividade esmagadora, o HCO3-contribui apenas 66% do comércio completo de CO2, enquanto que muito ainda este valor é de ∼85%.
Co2 Ligado Como Carbamato
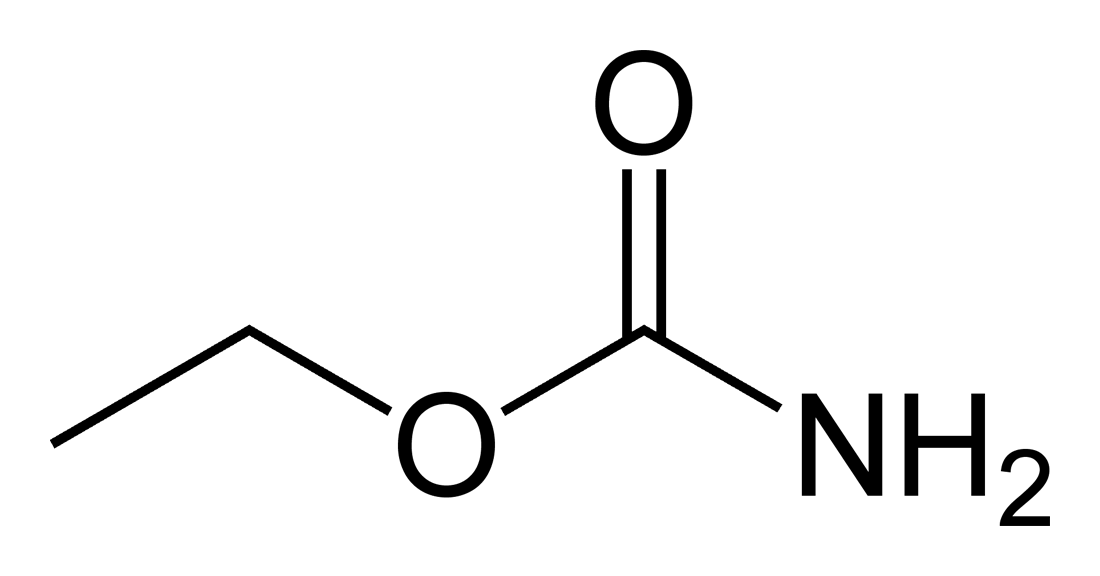
Carbamato
A medida do CO2 ligado como carbamato à hemoglobina em eritrócitos ou às proteínas plasmáticas baseia-se na imersão em O2 da hemoglobina e 2,3-difosfoglicérato (2,3-DPG), com foco nos eritrócitos, e na H+concentração por causa das plaquetas vermelhas e do plasma.
Durante a entrada de sangue através do músculo, a imersão de O2 e a fixação de H+ mudam extensivamente, especificamente durante o exercício. Seja como for, a expansão na dessaturação da hemoglobina e o incremento no foco de H+ experimentado pelas plaquetas vermelhas no estreito durante o exercício influenciam a medida do CO2 ligado à hemoglobina de forma inversa.
Embora a desoxigenação da hemoglobina construa a medida de CO2 ligado à hemoglobina, a fermentação diminui a medida de carbamato emoldurado pela hemoglobina.
Para calcular as fixações de carbamato dentro dos eritrócitos, utilizamos um arranjo solitário de constantes para o α-e β-correntes na condição oxigenada da hemoglobina e constantes separadas para o α-e β-correntes no estado desoxídico.
Como a estimativa de carbamato está sujeita à estimativa do equilíbrio de carbamato estável (pKc) e à harmonia de ionização consistente da recolha de aminoácidos (pKz) utilizada, erros na garantia destas constantes em várias investigações podem provocar alterações na contagem do carbamato.
Assim, utilizamos dois arranjos únicos de constantes para dar um indicador da flutuação das fixações de carbamato determinadas.